Мышечная механика, координация и энергетикаПотенциальные факторы человека состоят из скоростно-силовых качеств, координации движений, биоэнергетических возможностей и вегетативных систем обеспечения двигательной активности. Возможности реализации этого потенциала зависят от его физической, технической и психической подготовленности. Любой вид движений, в том числе во время занятий по физической подготовке происходит при помощи мышц, поэтому устройство мускулатуры и её строение необходимо хорошо знать всем занимающимся физкультурой, профессиональной физической подготовкой и спортсменам, У мужчин доля мышечной массы в теле может достигать до 50% от общего веса тела, у женщин - 35%. Значительно увеличить массу мышц в теле человека можно при помощи силовой тренировки. В тоже время недостаточная физическая активность влечёт за собой уменьшение процентной доли веса мышц и увеличению массы жира. А сейчас подробнее остановимся на рассмотрении устройства и работы мышц. Виды мышцВ человеческом организме есть скелетные и гладкие мышцы. Отдельное место занимает сердечная мышца. Скелетные (поперечнополостные) мышцы осуществляют движения и удерживают тело в равновесии. Эти мышцы укорачиваются при сокращении, и выполняют движения частей скелета посредством сухожилий. При интенсивной работе эти мышцы относительно быстро устают, и требуют достаточно длительного восстановления. Их работой человек способен управлять произвольно. Гладкие мышцы включены в состав внутренних органов человека. В них в результате взаимодействия сократительных элементов укорачиваются гладкомышечные клетки. Скорость их взаимного скольжения от 100 до 1000 раз меньше, чем в скелетных мышцах. Основная особенность гладких мышц заключается в том, что они хорошо приспособлены для длительного сокращения. При этом энергозатраты в них незначительны (приблизительно в 100-500 раз ниже, чем в скелетных), и соответственно их утомляемость также не высока. Несмотря на это, гладкие мышцы могут развивать такое же усилие, как и скелетные. Волевым усилием невозможно управлять работой гладких мышц без специальной подготовки. Сократительное усилие как скелетных, так и гладких мышц составляет 30 - 40 ньютон на один квадратный сантиметр поперечного сечения. Достоинства обеих групп мышц совмещает в себе сердечная мышца. Она может развивать большое усилие при малой утомляемости. Правильно организованная физическая подготовка развивает все группы мышц. Это оказывает положительное влияние не только на общую работоспособность, но и на работу внутренних органов. Мышцы преобразовывают химическую энергию в механическую работу, выделяя при этом тепловую энергию. Мышца состоит из мышечных клеток - миофибрилл, сосудов, соединительной ткани и нервов. В способных к сокращению миофибриллах вдоль мышечной клетки расположены тонкие и толстые нити - сократительные белки актин и миозин. Их суммарная массовая доля составляет 10% от массы всей мышцы. Нити актина и миозина имеют особую регулярную организацию - в них чередуются тёмные и светлые полосы. Поперечные тёмные перегородки - это так называемые Z-пластинки, которые разделяют миофибриллы на саркомеры, которые являются структурно - функциональными единицами сократительного аппарата мышц. В середине саркомера находятся несколько тысяч толстых миозиновых нитей, а на его концах около 2000 тонких актиновых нитей, которые закреплены на Z-пластинке как щетинки на щётке. Эта область саркомера называется I-диском, к которому со стороны, противоположной Z-пластинке прилегает более тёмная область - А-диск. В самой середине саркомера между А-дисками находится центральная Н-зона, которая не содержит актиновых нитей. В состоянии покоя I- и А-диски перекрываются незначительно. При сокращении мышцы актиновые нити начинают двигаться вдоль толстых нитей миозина к середине саркомера (рис. 1). При этом длина актиновых и миозиновых нитей остаётся неизменной, а изменяется лишь степень их перекрытия, которая при растяжении мышцы может уменьшиться до нуля. 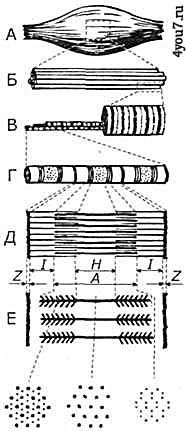
Рис. 1. Схематическое изображение строения мышцы.
А - мышца,
Б - пучок мышечных волокон,
В - мышечное волокно,
Г - миофибрилла,
Д - схематическое изображение саркомера, ограничено Z-линиями, с I-дисками, А-дисками и Н-зоной.
Е - поперечный срез на различных участках саркомера, дающий представление о распределении тонких и толстых нитей актина и миозина.
Нити миозина имеют поперечные выступы - так называемые мостики (головки), которые во время сокращения связывают миозиновую нить с актиновой (рис. 2). Наклоны этих головок создают усилие, благодаря которому нить актин продвигается к середине саркомера. При однократном таком движении поперечных мостиков саркомер сокращается приблизительно на 1% своей длины. За счёт того, что поперечные мостики выполняют серию движений, мышца может сократиться на 50% своей длины. Мышца может развивать большую силу благодаря суммации укорочений последовательно расположенных в миофибриллах саркомеров. 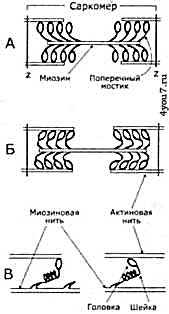
Рис. 2. Схематическое изображение поперечных мостиков саркомера.
Миозиновая нить с поперечными мостиками, соединенная с актиновыми нитями,
А - до,
Б – во время «гребцовых движений» (эти движения происходят асинхронно),
В - модель механизма генерации силы поперечными мостиками до (слева) и во время (справа) «гребкового движения»
Кроме того, благодаря эластичности поперечных мостиков мышца может развивать усилие и в статических условиях, то есть без изменения своей длины. Но даже в такой ситуации миозиновые «головки» не напряжены постоянно, через десятые или сотые доли секунды они отпускают миозиновые нити, после чего процесс прикрепления поперечного мостика к актиновой нити повторяется. Периодичность этого процесса составляет от 5 до 50 раз в секунду. Поскольку миозиновые «головки» прикрепляются и отсоединяются не одновременно, то количество прикреплённых головок в каждый момент времени примерно одинаково, и общее усилие, развиваемое мышцей остаётся постоянным. Скорость укорочения мышечного волокна и развиваемое им усилие возрастают во столько раз, во сколько длина миофибриллы больше длин составляющих его саркомеров. Этим объясняется тот факт, что длинная мышца сокращается быстрее, чем короткая. Сила развиваемая мышцей прямо пропорциональна количеству содержащихся в ней миофибрилл, иначе говоря, она тем сильнее, чем больше её поперечник. Структура скелетных мышцИз мышечных волокон состоят так называемые двигательные единицы (ДЕ), каждая из которых состоит из одного мотонейрона и группы иннервируемых им мышечных волокон (рис. 3). Типы мышц отличаются количеством ДЕ, которые, в свою очередь, тоже очень сильно отличаются размерами. Так один мотонейрон может иннервировать от нескольких ДЕ до 2000. При этом каждая мышца содержит ДЕ с разным количеством волокон. Мышцы, предназначенные для выполнения точной работы, содержат от 1500 до 3000 ДЕ, которые, в свою очередь, содержат от 8 до 50 миофибрилл. В то же время, мышцы, предназначенные для выполнения менее точных движений, но с большим усилием, содержат значительно меньшее количество ДЕ, которые содержат значительно больше волокон - от 600 до 2000. 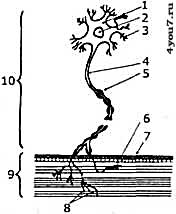
Рис. 3. Схема строения двигательной единицы.
1 - тело мотонейрона,
2 – ядро,
3 - дендриты - короткие отростки,
4 - аксон - длинный отросток нервного волокна,
5 - оболочка нервного волокна,
6 - концевые веточки аксона,
7 - нервно-мышечные синапсы, через которые мышце передаются нервные импульсы,
8 - мышечное волокно с миофибриллами,
9 - оболочка мышечного волокна.
ДЕ состоит из двух основных типов волокон. Первый тип - это сокращающиеся быстро, и развивающие при этом большое усилие (Fast twitch - FT) волокна. Второй тип - это менее быстрые и сильные, но значительно более выносливые (Slow twitch - ST) волокна. Быстрые мышечные волокна содержат повышенное количество гликогена, и имеют высокую активность анаэробных гликолитических ферментов. Использование в этих волокнах внутримышечных энергетических субстратов не обеспечивает им запас выносливости, из-за чего эти волокна эффективно выполняют свою функцию с максимальной энергоотдачей в течение не более четырёх минут. Этот тип волокон иначе называют Fast glycolysis (FG) по международной классификации. Для выполнения своих функций в течение значительно большего времени, но, развивая при этом меньшее усилие, лучше приспособлены медленные волокна. Такими свойствами они обладают за счёт аэробного, иначе говоря, окислительного способа функционирования. Эти волокна обладают хорошо развитой капиллярной сетью, которая активно снабжает их кислородом из крови, чем и обеспечивается их продолжительная работа. В ST - волокнах содержится также повышенное количество миоглобина и митохондрий, в которых протекают процессы окисления. Кроме того, в этих волокнах запасено повышенное, по сравнения с FT - волокнами, количество жиров в виде триглицеридов, которые являются субстратами окисления. Этот тип волокон иначе называется Slow oxydative (SO) в соответствии с международной классификацией. Среди быстрых волокон выделяют подтип окислительно-гликолитических (FOG - тип), которые способны выполнять работу с большой интенсивностью в аэробном (окислительном) режиме, в тоже время с мощным гликолитическим энергообразованием. Такой тип волокон считается промежуточным между FG и SO - типами. Соотношение волокон, составляющих мышцы не изменяется в течение жизни, поскольку оно определено генетически, но при помощи соответствующей тренировки можно увеличить толщину волокон всех типов. Регуляция напряжения мышцВсе движения в человеческом теле происходят за счёт сокращения соответствующих мышц. Этим процессом управляет нервная система. Степень напряжения сокращающихся мышц регулируется следующими способами: - при помощи изменения количества активных в данный момент ДЕ;
- частотой импульсации мотонейронов;
- временной связью активности ДЕ.
ДЕ активизируются за счёт подачи её мотонейроном импульса мышечным волокнам. При этом на данный импульс реагируют все волокна, входящие в состав этой ДЕ. Каждая ДЕ сокращается с силой, прямо пропорциональной количеству входящих в её состав мышечных волокон. При этом усилие, развиваемое разными ДЕ колеблются от нескольких миллиньютон до нескольких ньютон. На практике, при выполнении любого движения, задействуется некоторое количество ДЕ, поскольку возможности одной ДЕ невелики. Такое явление в физиологии называется пространственной суммацией. Количество задействованных в каждый момент ДЕ определяется управляющими импульсами, поступающими от нервной системы мотонейронам мышц, причём меньшие по размерам мотонейроны имеют меньший порог «срабатывания», то есть они реагируют на менее сильные управляющие импульсы. Размер мотонейрона, в свою очередь, зависит от размеров ДЕ, поэтому малые мышечные напряжения обеспечиваются при помощи малых и медленных ДЕ. Сокращение ДЕ мышц происходит не от одиночных импульсов, а от их серий. Частота управляющих импульсов в таких сериях лежит в пределах от 5 импульсов в секунду до 50. Когда каждый последующий
импульс в серии подаётся до того момента, как ДЕ успела расслабиться от воздействия предыдущего, происходит нарастание развиваемого этой ДЕ усилия. Такое сокращение мышцы называется тетаническим, и его продолжительность может в тысячи раз превышать длительность сокращения мышцы от одиночного импульса (рис. 4). Регулировка силы сокращения мышцы таким методом называется «временной суммацией». 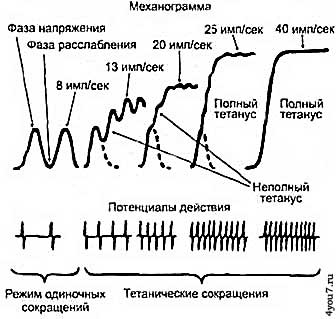
Рис 4. Режимы сокращений двигательной единицы импульсами различной частоты.
При постепенном увеличении напряжения в работу включается всё более крупные ДЕ. При подаче 20 нервных импульсов в секунду (20 Гц) потенциал выносливых SO - волокон может быть полностью исчерпан, в то время, как частота нервных импульсов 25 - 30 Гц приводит лишь к умеренному напряжению FG - волокон, а для их максимального напряжения требуется частота импульсов 45-50 Гц. Мышца, в которой большинство составляющих её волокон относятся к SO - типу, может значительно дольше поддерживать напряжение, хотя и с меньшим усилием, чем мышца, состоящая преимущественно из волокон FG - типа. Таким образом, частота управляющих импульсов, необходимая для полного тетанического сокращения ДЕ зависит от мышечных волокон, входящих в её состав. При выполнении скелетными мышцами движений, не требующих больших усилий работают низкопороговые ДЕ, причем не одновременно, а каждая из них активируется в разные моменты времени так, что общее усилие, развиваемое мышцей остаётся постоянным во времени. При мышечной работе средней интенсивности длительное время по мере утомления низкопороговых мышц в работу начинают включаться всё более крупные и более высокопороговые ДЕ. При такой работе внутримышечный источник энергии гликоген сначала расходуется в SO - волокнах, а затем и в FG - волокнах. При преодолении мышцами высокого внешнего сопротивления с большой скоростью работают в основном высокопороговые ДЕ с волокнами FG - типа, которые активируются высокочастотными импульсами, которые следуют сериями. При этом в паузах между «пачками» нервных управляющих импульсов мышца продолжает оставаться в напряжении. В таком режиме работы мышцы в случае увеличения внешнего сопротивления, для его преодоления будет увеличиваться длительность «пачек» управляющих нервных импульсов. Внутримышечная координацияВнутримышечной координацией называется способность человека синхронно управлять работой своих ДЕ. Такое умение очень важно по следующей причине: при выполнении обычных действий нетренированный человек способен одновременно активировать не более 60% ДЕ своих мышц, в то время как тренированный спортсмен в момент развития максимальной силы способен активировать до 85% ДЕ в соответствующих мышцах. Развиваемая мышцей сила тяги увеличивается при выполнении динамической работы уступающего характера, иначе говоря, при принудительном растягивании мышц. Причём увеличение силы составляет 10% у спортсменов и до 35% у нетренированных людей. Такая прибавка силы называется непроизвольно мобилизованной силой. Разница между произвольно и непроизвольно мобилизованной силой называется дефицитом силы. Величина развиваемой силы возрастает с увеличением скорости растягивания, а в преодолевающем режиме работы с увеличением скорости развиваемая сила снижается. Большой дефицит силы вызван низкой внутримышечной координацией, улучшая которую можно добиться существенного прироста силы. В тоже время, если дефицит силы не более 10%, то для увеличения развиваемого
мышцами усилия необходимо наращивать мышечную массу. Мышечная механикаДля понимания механических свойств весьма сложно устроенной мышцы, в большинстве случаев, достаточно воспользоваться её упрощенной моделью и рассматривать мышцы как комбинацию упругих и сократительных элементов (рис. 5). При этом последними являются те участки саркомеров, на которых актиновые и миозиновые нити друг друга перекрывают, именно здесь и происходит генерация мышечного усилия. 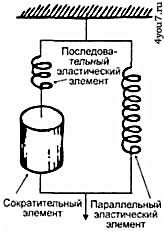
Рис 5. Аналоговая модель строения мышцы.
СЭ - сократительный элемент,
ПаЭЭ - параллельный эластический элемент,
ПоЭЭ - последовательный эластический элемент.
Расслабленная мышца оказывает очень низкое сопротивление растяжению, поскольку актиновые и миозиновые нити очень легко скользят друг относительно друга. Поэтому укоротившуюся в результате сжатия мышцу очень легко растянуть внешним воздействием. При этом покоящуюся мышцу можно растянуть и сверх её длины, однако уже с приложением большего усилия, когда будут растягиваться её эластичные структуры. К таким элементам относится оболочка вокруг мышечного волокна (сарколемма), элементы соединительной ткани между волокнами и их пучками и другие элементы мышечных клеток. Сила, развиваемая мышцей, зависит от уровня её предварительного растяжения, из-за которого зависит степень перекрытия актина и миозина в саркомере (рис. 6). Максимальное усилие развивается при длине саркомера 2 - 2,2 мкм. С уменьшением его длины снижается усилие, развиваемое мышцей, поскольку актиновые и миозиновые нити начинают мешать друг другу. Когда сокращение длины мышцы достигает 50 - 70%, она прекращает развивать усилие. 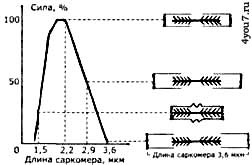
Рис. 6. Зависимость между силой сокращения, длиной саркомера и степенью перекрывания актиновых и миозиновых нитей.
А - зависимость между максимальной изометрической силой, развиваемой саркомером во время тетануса и его длиной;
Б - взаимное перекрывание актиновых и миозиновых нитей при различной длине саркомера.
Сила, развиваемая, мышцей снижается и тогда, когда мышца растягивается больше, чем её длина в покое ввиду того, что нити актина вытягиваются из пучка нитей миозина, и степень их взаимного перекрытия начинает уменьшаться. Она снижается до 50% при длине саркомера 2,9 мкм, а при его длине 3,6 мкм актиновые и миозиновые нити уже не пересекаются, и мышца не может развивать усилие. На своих концах мышцы переходят в упругие сухожилия, посредством которых усилие передаётся от этих мышц на костные рычаги. При резком сокращении или внезапном сильном внешнем воздействии сухожилия несколько растягиваются, смягчая тем самым нагрузку на мышцы. Прочность мышечной ткани на растяжение 60 Н/см2, а сухожилия -700 Н/см2. Места переходов мышц в сухожилия - это наиболее часто травмируемые участки, поэтому перед каждым занятием по развитию мышечной силы необходима основательная разминка мышц. Ненагруженная мышца сокращается с максимальной скоростью, а по мере увеличения нагрузки на неё, скорость её сокращения будет снижаться. При быстром сокращении мышца способна развивать меньшее усилие, чем при медленном (рис. 7). При помощи активации большего или меньшего количества ДЕ (при этом меняется относительная нагрузка на каждое мышечное волокно), человек способен регулировать скорость сокращения мышц при непредельных нагрузках. 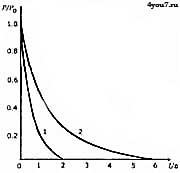
Рис 7. Зависимость между относительной силой (Р/Ро) и максимальной скоростью сокращения (Iо/с) для мышц, состоящих из волокон ST- (1) и FT- (2) типа.
Межмышечная координацияНикакое движение в человеческом теле не выполняется при помощи только одной мышцы, а всегда задействуется группа мышц. Мышцы и их группы выполняют два вида функций, которые невозможно чётко разграничить между собой - это движение и поддержание нужной позы. Для правильного выполнения обеих функций работающие мышцы должны быть очень хорошо скоординированы между собой в пространственно-временных и динамико-временных отношениях. Мышцы, которые согласованно выполняют какое либо движение называются агонистами, а те мышцы, которые выполняют движение в противоположном направлении - антагонистами. При сгибании конечностей агонисты сокращаются, а антагонисты расслабляются, что приводит к дополнительному ускорению движения в самом начале выполнения движения, после чего наблюдается снижение скорости движения, в конце которого напряжение антагонистов его останавливает. Согласованная работа агонистов и антагонистов в большой мере обусловлена хорошей растянутостью мышц, которая позволяет выполнять движения с большей амплитудой, и, соответственно, с большим путём приложения усилия. При выполнении циклических движений часть энергии может накапливаться в эластических структурах мышц и связок за счёт их упругой деформации, и высвобождаться на обратном движении. Такой режим работы мышцы называется рекуперацией энергии, вклад которой в общую работу возрастает с увеличением скорости выполнения движений. Роль хорошей межмышечной координации возрастает при выполнении сложных движений, в которых участвует большое количество мышц. Её развитие позволит в таких случаях увеличить развиваемую силу, скорость, гибкость и выносливость. Обмен веществВ основе жизнедеятельности всех живых организмов лежит обмен веществ между ними и окружающей их средой (метаболизм). Этот процесс химического обновления живых организмов идёт постоянно и регулируется самостоятельно. Цель обмена веществ заключается в том, чтобы обеспечить постоянство состава и внутренних параметров организма, рост, способность выполнять движения и адаптироваться при изменениях в окружающей среде. Метаболизм состоит из двух необходимых составляющих: в процессе обмена веществ их усвоение и биологическое превращение называется ассимиляция, а распад веществ и их выведение - диссимиляция. Для всех процессов, проходящих в живом организме, таких как синтез необходимых веществ, сокращение мышц и управление ими посредством нервных импульсов, требуется энергия. Она получается в результате диссимиляции, то есть биологического окисления веществ, служащих её источником. Такие вещества организм получает с пищей. Из неё же синтезируется аденозинтрифосфат (АТФ), который выполняет роль непосредственного источника энергии для выполнения всех биологических функций организма, в том числе и для работы мышц. В таблице 1 приведено распределение средних суточных затрат энергии человеческого организма. В состоянии покоя энергозатраты человека составляют приблизительно 5,25 кДж/мин (1,25 ккал/мин). При физических нагрузках затраты энергии человеком возрастают в 10 - 12 раз. Таблица 1
Функция | Энергозатраты, кДж | Синтез АТФ | 3770 | Синтез белков, жиров и углеводов | 1740 | Поддержание ионных градиентов | 900 | Работа кардиореспираторной системы | 1130 | Всего | 7540 |
На протяжении жизни человека интенсивность обменных процессов не одинакова. Так, сразу после рождения скорость синтеза веществ превышает скорость их распада, затем, приблизительно к 17 - 19 годам скорости этих процессов становятся равными, и рост организма прекращается. В старости скорость диссимиляции преобладает над скоростью ассимиляции, из-за чего снижается скорость биосинтеза многих важных веществ. Некоторого преобладания процесса ассимиляции над диссимиляцией можно достичь при помощи физической тренировки. Это приведёт к накоплению сократительных белков, ферментов и энергетических субстратов. Величиной и направленностью физической нагрузки можно регулировать качественный и количественный состав этих накоплений. При физической нагрузке усиливаются энергозатраты, и следовательно, возникающий энергетический дефицит затормаживает процессы ассимиляции, что приводит к преобладанию процессов диссимиляции. После снижения физической нагрузки для восполнения затраченных энергоресурсов наступает преобладание процессов ассимиляции. При этом возрастает общая интенсивность обменных процессов, что создаёт условия для быстрого обновления тканей человеческого организма. Такая ситуация в детстве и юности способствует усиленному росту, а в другие периоды жизни стимулирует повышенную функциональную активность. Из вышесказанного можно сделать вывод о безусловной пользе физических тренировок на протяжении всей жизни человека.
| 











